ქოლესტერინის სინთეზის რეგულირება
ქოლესტერინის სინთეზის რეგულირება - მისი საკვანძო ფერმენტი (HMG-CoA რედუქტაზა) ხორციელდება სხვადასხვა გზით.
HMG რედუქტაზას ფოსფორილირება / დეფოსფორილირება. ინსულინ / გლუკაგონის თანაფარდობის მატებით, ეს ფერმენტი ფოსფორილდება და გადადის აქტიურ მდგომარეობაში. ინსულინის მოქმედება ხორციელდება 2 ფერმენტის მეშვეობით.
HMG-CoA ამცირებს კინაზას ფოსფატაზას, რომელიც კინაზას გადააქცევს არააქტიურ დეფოსფორილირებულ მდგომარეობაში:
ფოსფოტაზა HMG-CoA რედუქტაზა მისი დეფოსფორილირებული აქტიურ მდგომარეობაში გადაკეთებით. ამ რეაქციების შედეგია HMG-CoA რედუქტაზის დეფოსფორილირებული აქტიური ფორმის წარმოქმნა.
შესაბამისად, შთანთქმის პერიოდში, ქოლესტერინი იზრდება. ამ პერიოდის განმავლობაში, საწყისი სუბსტრატის არსებობა ხდება ქოლესტერინის - აცეტილ - CoA სინთეზისთვის (ნახშირწყლებისა და ცხიმების შემცველი საკვების ჭამის შედეგად, რადგან გლუკოზისა და ცხიმოვანი მჟავების დაშლის დროს წარმოიქმნება აცეტილ CoA).
პოსტაბსორბენტულ მდგომარეობაში გლუკაგონი A პროტეგენგენაზას საშუალებით ასტიმულირებს HMG - CoA - ფოსფორილირებას - რედუქტაზას, გადააქცევს მას არააქტიურ მდგომარეობაში. ეს მოქმედება გაძლიერებულია იმით, რომ ამავე დროს გლუკაგონი ასტიმულირებს HMG-CoA რედუქტაზას ფოსფოტაზას ფოსფორილირებას და ინაქტივირებას, რითაც HMG-CoA რედუქტაზას ინარჩუნებს ფოსფორილირებულ არააქტიურ მდგომარეობაში. შედეგად, პოსტბსორბციის პერიოდში და მარხვის პერიოდში ქოლესტერინის სინთეზი ინჰიბირდება ენდოგენური სინთეზით. თუ საკვებში ქოლესტერინის შემცველობა 2% -მდე იქნა მიღებული, მაშინ ენდოგენური ქოლესტერინის სინთეზი მკვეთრად შემცირდა. მაგრამ ქოლესტერინის სინთეზის სრული შეწყვეტა არ ხდება.
საკვებიდან მომავალი ქოლესტერინის გავლენის ქვეშ ქოლესტერინის ბიოსინთეზის ინჰიბირების ხარისხი განსხვავდება პიროვნებიდან. ეს მიუთითებს ქოლესტერინის წარმოქმნის პროცესების ინდივიდუალურობაზე. ქოლესტერინის სინთეზის ინტენსივობის შემცირებით შესაძლებელია სისხლში მისი კონცენტრაციის შემცირება.
თუ ერთის მხრივ ორგანიზმში ქოლესტერინის საკვების მიღებასა და მასში სინთეზს შორის წონასწორობა და მეორეს მხრივ, ნაღვლის მჟავების და ქოლესტერინის ექსკრეცია დარღვეულია, ქსოვილებში ქოლესტერინის კონცენტრაცია და სისხლი იცვლება. ყველაზე სერიოზული შედეგები უკავშირდება სისხლში ქოლესტერინის კონცენტრაციის ზრდას (ჰიპერქოლესტერინემია), ხოლო ათეროსკლეროზისა და ქოლელითაზიის განვითარების ალბათობა იზრდება.
ოჯახური ჰიპერქოლესტერინემია (HCS) - ეს ფორმა გაცილებით გავრცელებულია - 200 ადამიანზე დაახლოებით 1 პაციენტი. HCS- ში მემკვიდრეობითი დეფექტი არის უჯრედების მიერ LDL შეწოვის დარღვევა და, შესაბამისად, LDL კატაბოლიზმის სიჩქარის დაქვეითება. შედეგად, სისხლში LDL- ის კონცენტრაცია იზრდება, ისევე როგორც ქოლესტერინი, რადგან მასში ბევრია LDL- ში. ამიტომ, HCS– ით, ქოლესტერინის დეპონირება ხდება ქსოვილებში, კერძოდ კანში (ქსანთომა), არტერიების კედლებში.
HMG-CoA რედუქტაზას სინთეზის ინჰიბირება
ქოლესტერინის მეტაბოლური გზის საბოლოო პროდუქტი. ეს ამცირებს HMG-CoA რედუქტაზას გენის ტრანსკრიპციის სიჩქარეს, რითაც ახდენს საკუთარი სინთეზის შეფერხებას. ღვიძლი აქტიურად ახდენს ნაღვლის მჟავების სინთეზს ქოლესტერინისგან, და ამიტომ ნაღვლის მჟავები აფერხებენ HMG-CoA რედუქტაზის გენის მოქმედებას. ვინაიდან HMG-CoA რედუქტაზა არსებობს დაახლოებით 3-ის სინთეზის შემდეგ, ამ ქოლესტერინის ფერმენტის სინთეზის დათრგუნვა ეფექტური რეგულირებაა.
ვერ იპოვნეთ ის, რასაც ეძებდით? გამოიყენეთ ძებნა:
ქოლესტერინის ეთერის გაცვლა
ქოლესტერინის ფონდი შეიცავს უფასო ქოლესტერინის და ქოლესტერინის ეთერებს, რომლებიც გვხვდება როგორც უჯრედებში, ასევე სისხლის ლიპოპროტეინებში.
ნაწილი II მეტაბოლიზმი და ენერგია
უჯრედებში ქოლესტერინის ეთერიფიკაცია ხდება აცილ-კოა-ქოლესტერინის-აცილტრანსფერაზას (AChAT) მოქმედებით.
აცილ-კოა + ქოლესტერინი - * HS-KoA + აციკლეოლესტერინი
ადამიანის უჯრედებში, ძირითადად, ლინოლეიქოლესტერინი ვითარდება. თავისუფალი ქოლესტერინისგან განსხვავებით, უჯრედულ მემბრანებში მისი ესტერი შეიცავს ძალიან მცირე რაოდენობით და ძირითადად გვხვდება ციტოზოლში, როგორც ლიპიდური წვეთების ნაწილი. ეთერების წარმოქმნა შეიძლება ჩაითვალოს, ერთი მხრივ, როგორც მემბრანებიდან ჭარბი ქოლესტერინის მოცილების მექანიზმი, და, მეორე მხრივ, როგორც უჯრედში ქოლესტერინის შენახვის მექანიზმი. რესურსების მობილიზაცია ხდება ესტერაზას ფერმენტების მონაწილეობით, რომლებიც ატენიანებენ ქოლესტერინის ეთერებს:
აცილქოლესტერინი + H 2O - * ცხიმოვანი მჟავა + ქოლესტერინი
ესტერების სინთეზი და ჰიდროლიზი ხდება ბევრ უჯრედში, მაგრამ განსაკუთრებით აქტიურია თირკმელზედა ჯირკვლის ქერქის უჯრედებში: ამ უჯრედებში ყველა ქოლესტერინის 80% -მდე წარმოდგენილია ესტერები, ხოლო სხვა უჯრედებში, ჩვეულებრივ, 20% -ზე ნაკლები.
სისხლში ლიპონპროტეინებში ესთერების წარმოქმნა ხდება ლეციტინ-ქოლესტერინის აცილტრანსფერაზას (LHAT) მონაწილეობით, რომელიც კატალიზაციას ახდენს აცილის ნარჩენების გადატანას ლეციტინის i- პოზიციიდან ქოლესტერინამდე (ნახ. 10.31). LHAT წარმოიქმნება ღვიძლში, გამოიყოფა სისხლში და თან ერთვის ღვიძლში. სხვადასხვა ლიპოპროტეინების ეთერიფიკაციის სიჩქარე მნიშვნელოვნად განსხვავდება და დამოკიდებულია აპოლიპოპროტეინების არსებობაზე, რომლებიც ააქტიურებენ LHAT- ს (ძირითადად აპი-ატ, აგრეთვე CI) ან ამ ფერმენტის ინჰიბირებას (C-II) .იგი ყველაზე აქტიური LHAT in HDL, რომელშიც ა უნდა Olee 2/3 ყველა ცილა. ყველაზე coli პატივით ჩამოყალიბდა ეთერები oleic და linoleic მჟავები. სხვა ლიპოპროტეინების ester ფორმირება ხდება უფრო ნელა, ვიდრე HDL.
სურ. 10.31. ქოლესტერინის ეთერების წარმოქმნა LHAT მოქმედების ქვეშ
LHAT ლოკალიზებულია HDL- ის ზედაპირულ ფენაში და იყენებს ქოლესტერინს ფოსფოლიპიდურ მონოლიტერში, როგორც სუბსტრატს. აქ ჩამოყალიბებული ქოლესტერინის ესტერიები, მათი სრული ჰიდროფობიულობის გამო, ცუდად ინარჩუნებენ
თავი 10. მეტაბოლიზმი და ლიპიდური ფუნქცია
ფოსფოლიპიდური მონოლიტერი და ჩაეფლო ლიპოპროტეინების ცხიმის ბირთვში. ამავდროულად, ქოლესტერინის ადგილი თავისუფლდება ფოსფოლიპიდურ მონოლიტერში, რომელიც ქოლესტერინით შეიძლება შეივსოს უჯრედული მემბრანებიდან ან სხვა ლიპოპროტეინებისგან. ამრიგად, HDL, როგორც ჩანს, ქოლესტერინის ხაფანგია LHAT– ის მოქმედების შედეგად.
ნაღვლის მჟავების სინთეზი
ღვიძლში, ქოლესტერინის ნაწილი გარდაიქმნება ნაღვლის მჟავებად. ნაღვლის მჟავები შეიძლება ჩაითვალოს ქოლანმჟავას წარმოებულებად (ნახ. 10.32).
ქოლანის მჟავა, როგორც ასეთი, სხეულში არ იქმნება. ჰეპატოციტების დროს, ქოლესტერინი პირდაპირ წარმოქმნის ცენოდეოქსიქოლიურ და ქოლინის მჟავებს - ნაღვლის პირველადი მჟავებს (ნახ. 10.33, იხ. აგრეთვე ნახ. 10.12).
ქოლესტერინის ბიოსინთეზი
ქოლესტერინის ბიოსინთეზი ხდება ენდოპლაზმურ რეტიკულუმში. მოლეკულში ნახშირბადის ყველა ატომის წყაროა აცეტილ-SCoA, რომელიც აქ მოდის მიტოქონდრიიდან, როგორც ციტრატის ნაწილი, ისევე როგორც ცხიმოვანი მჟავების სინთეზში. ქოლესტერინის ბიოსინთეზი მოიხმარს 18 ატფ მოლეკულას და 13 NADPH მოლეკულს.
ქოლესტერინის ფორმირება ხდება 30-ზე მეტ რეაქციაში, რომელთა დაჯგუფება შესაძლებელია რამდენიმე ეტაპზე.
1. მეგალონური მჟავის სინთეზი.
პირველი ორი სინთეზის რეაქცია ემთხვევა კეტოგენეზის რეაქციებს, მაგრამ 3-ჰიდროქსი-3-მეთილგლუტარილ-ScoA სინთეზის შემდეგ, ფერმენტი შემოდის ჰიდროქსიმეთილ-გლუტარილ-ScoA რედუქტაზა (HMG-SCOA რედუქტაზა), წარმოქმნის მივალონური მჟავა.
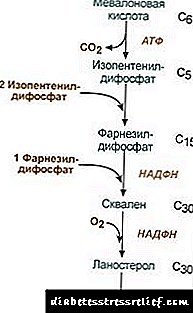 |
ქოლესტერინის სინთეზის რეაქციის სქემა2. იზოპენტენილის დიფოსფატის სინთეზი. ამ ეტაპზე, ფოსფატის სამი ნარჩენი მიმაგრებულია მევალონიუმის მჟავას, შემდეგ იგი ხდება დეკარბოქსილირებულ და დეჰიდროგენებად. 3. იზოპენტენილის დიფოსფატის სამი მოლეკულის შეთავსების შემდეგ, სინთეზირებულია ფარნესილის დიფოსფატი. 4. სკალელის სინთეზი ხდება მაშინ, როდესაც შეკრულია ორი ფარნესილის დიფოსფატის ნარჩენები. 5. რთული რეაქციების შემდეგ, წრფივი სკალლენი ციკლირდება ლანოსტეროლამდე. 6. მეტილის ჯგუფების ჭარბი მოცილება, მოლეკულის აღდგენა და იზომერიზაცია იწვევს ქოლესტერინის გამოვლენას. ჰიდროქსიმეთილგლუტარილ-S-CoA რედუქტაზას საქმიანობის რეგულირება3. ქოლესტერინის ბიოსინთეზის სიჩქარე ასევე დამოკიდებულია კონცენტრაციაზე სპეციფიკური გადამზიდავი ცილაჰიდროფობიური შუალედური სინთეზის მეტაბოლიტების დაკავშირება და ტრანსპორტირება. შეგიძლიათ მოსთხოვოთ ან დატოვოთ თქვენი აზრი. რეგულირების მთავარი წერტილი არის მეგალონის მჟავის წარმოქმნის რეაქცია. 1. ალლოსტერიული რეგულირება. ქოლესტერინი, და ღვიძლში - და ნაღვლის მჟავები აჩერებენ HMG-CoA რედუქტაზას. 2. HMG-CoA რედუქტაზას ქოლესტერინის სინთეზის რეპრესიები. 3. რეგულირება HMG-CoA რედუქტაზის ფოსფორილირება-დეფოსფორილირების გზით, აქტიური არაფოსფორილირებული ფორმა. გლუკაგონი იწვევს დეაქტივაციას, ხოლო ინსულინი იწვევს აქტივაციას რეაქციების რთული კასკადის საშუალებით. ამრიგად, ქოლესტერინის სინთეზის სიჩქარე იცვლება შთამნთქმელი და პოსტაბსორბციული მდგომარეობების ცვლილებით. 4. ღვიძლში HMG-CoA რედუქტაზას სინთეზის მაჩვენებელი ექვემდებარება მკაფიო დიარეალურ რყევებს: მაქსიმუმ შუაღამისას, ხოლო მინიმუმით დილით. ქოლესტერინის ეთერის გაცვლა უჯრედებში ქოლესტერინის ეთერიფიკაცია ხდება ექსპოზიციის დროს აცილ-კოა-ქოლესტერინი-აცილტრანსფერაზა (AHAT): Acyl-CoA + ქოლესტერინი ® NS-CoA + აციკლეოლესტერინი უჯრედებში, ძირითადად, ლინოლეიქოლესტერინი ვითარდება. ესტრები ძირითადად გვხვდება ციტოზოლში, როგორც ლიპიდური წვეთების ნაწილი. ესტერების წარმოქმნა შეიძლება ჩაითვალოს, ერთი მხრივ, როგორც მემბრანებიდან ჭარბი ქოლესტერინის მოცილების მექანიზმი, და, მეორე მხრივ, როგორც უჯრედში ქოლესტერინის შენახვის მექანიზმი. რეზერვების მობილიზაცია ხდება ფერმენტების მონაწილეობით esteraseქოლესტერინის ესტერინების ჰიდროლიზაცია: ასიქსოლესტერინი + H2O ® ცხიმოვანი მჟავა + ქოლესტერინი ესტერების სინთეზი და ჰიდროლიზი განსაკუთრებით აქტიურია თირკმელზედა ჯირკვლის ქერქის უჯრედებში. სისხლში ლიპოპროტეინებში ester ფორმირება ხდება მონაწილეობით ლეციტინი-ქოლესტერინი-აცილტრანსფერაზა (LHAT), რომელიც ახდენს აცილის ნარჩენების ლეციტინიდან ქოლესტერინში გადატანას. LHAT იქმნება ღვიძლში, გამოიყოფა სისხლში და მიმაგრებულია ლიპოპროტეინებით. ყველაზე აქტიური LHAT HDL- ში, სადაც ის ლოკალიზებულია ზედაპირის ფენაში. აქ წარმოქმნილი ქოლესტერინის ესტერები ჰიდროფობიურია და ლიპიდურ ბირთვში ჩაეფლო. ფოსფოლიპიდურ მონოლიტერში, ქოლესტერინისთვის თავისუფალი სივრცეა, რომელიც ქოლესტერინით შეიძლება შეივსოს უჯრედული მემბრანებიდან ან სხვა ლიპოპროტეინებისგან. ამრიგად, HDL, როგორც ჩანს, ქოლესტერინის ხაფანგია LHAT– ის მოქმედების შედეგად. ნაღვლის მჟავების სინთეზი ღვიძლში, ქოლესტერინის ნაწილი გარდაიქმნება ნაღვლის მჟავებად. ნაღვლის მჟავები შეიძლება ჩაითვალოს ქოლანმჟავას წარმოებულებად. ქოლანის მჟავა, როგორც ასეთი, სხეულში არ იქმნება. ქოლესტერინისგან ჰეპატოციდებში წარმოიქმნება პირველადი ნაღვლის მჟავები - ჩენოდოქსიქოლიური და ქოლელი. ნაწლავის ფლორის ფერმენტების მოქმედების შედეგად ნაწლავში ნაღვლის სეკრეციის შემდეგ, მათგან იქმნება მეორადი ნაღვლის მჟავები - ლითოქოლური და დეოქსიქოლიური. ისინი ნაწლავებიდან შეიწოვება, პორტატული ვენის სისხლით შედის ღვიძლში, შემდეგ კი ნაღვლის ღრუში. ნაღვლის შემადგენლობაში შედის ძირითადად კონიუგირებული ნაღვლის მჟავები, ანუ მათი ნაერთები გლიცინით ან ტაურინით. ნაღვლის მჟავების კონცენტრაცია ნაღველში არის დაახლოებით 1%. ნაღვლის მჟავების ძირითადი ნაწილი მონაწილეობს ჰეპატოცენტრული მიმოქცევა.ნაღვლის მჟავების მცირე ნაწილი - დაახლოებით 0.5 გ დღეში - გამოიყოფა განავლით. ეს ანაზღაურდება ღვიძლში ახალი ნაღვლის მჟავების სინთეზით, ნაღვლის მჟავის ფონდი განახლდება დაახლოებით 10 დღეში. ქოლესტერინი ასევე გამოიყოფა ძირითადად ნაწლავებით. იგი ნაწლავებში შედის საკვებით და ღვიძლიდან, როგორც ნაღვლის ნაწილი. სისხლში შეიწოვება ქოლესტერინი შეიცავს ნაღვლისგან წარმოქმნილ ნაწილს (ენდოგენური ქოლესტერინიღვიძლში სინთეზირებულია) და საკვებიდან მიღებული წილი (ეგზოგენური ქოლესტერინი) ქსოვილებში ქოლესტერინის მოცილება ხდება მისი დაჟანგვით ღვიძლში მჟავა მჟავების გამო, რასაც მოჰყვება განავლით მათი განდევნა (დღეში დაახლოებით 0,5 გ დღეში) და უცვლელი ქოლესტერინის (აგრეთვე განავლით) ექსკრეციით. სტაციონარულ მდგომარეობაში: (ქოლესტერინიდასასრული + ქოლესტერინიყოფილი) - (ქოლესტერინიექსკრე + ნაღვლის მჟავებიექსკრე) = 0 თუ ეს ბალანსი დარღვეულია, ცვლის ქოლესტერინის კონცენტრაცია ქსოვილებში და სისხლში. სისხლში ქოლესტერინის მომატება - ჰიპერქოლესტერინემია. ეს ზრდის ათეროსკლეროზისა და ნაღვლის ბუშტის დაავადების ალბათობას. ლიპიდების გაცვლის რეგულირება ლიპიდების მეტაბოლიზმი რეგულირდება ცენტრალური ნერვული სისტემის მიერ. გრძელი უარყოფითი ემოციური სტრესი, კატეხოლამინების სისხლში სისხლძარღვში განთავისუფლების გაზრდამ შეიძლება გამოიწვიოს შესამჩნევი წონის დაკარგვა. მოქმედება გლუკაგონი ლიპოლიზურ სისტემაზე მსგავსია კატექოლამინების მოქმედება. ადრენალინი და ნორეპინეფრინი ქსოვილოვანი ლიპაზის აქტივობის გაზრდა და ცხიმოვან ქსოვილში ლიპოლიზის სიჩქარე, შედეგად იზრდება სისხლის პლაზმაში ცხიმოვანი მჟავების შემცველობა. ინსულინი აქვს ადრენალინისა და გლუკაგონის საპირისპირო მოქმედება ლიპოლიზზე და ცხიმოვანი მჟავების მობილიზაციაზე. ზრდის ჰორმონი ასტიმულირებს ლიპოლიზს, ახდენს აკენილატ ციკლაზას სინთეზს. ჰიპოფიზის ჰიპოფიზი იწვევს ორგანიზმში ცხიმის დეპონირებას (ჰიპოფიზის სიმსუქნე). თიროქსინი, სქესის ჰორმონებიასევე გავლენას ახდენს ლიპიდების მეტაბოლიზმზე. სქესობრივი ჯირკვლების მოცილება ცხოველებში იწვევს ზედმეტი ცხიმის დეპონირებას. LIPID– ის გაცვლა ქოლესტერინი არის სტეროიდი, რომელიც სპეციფიკურია ცხოველების ორგანიზმებისთვის. ადამიანის ორგანიზმში მისი წარმოქმნის მთავარი ადგილი არის ღვიძლი, რომელშიც სინთეზირდება ქოლესტერინის 50%, წვრილი ნაწლავის შემადგენლობაში 15-20%, დანარჩენი სინთეზირებულია კანში, თირკმელზედა ჯირკვლის ქერქში და გონებში. ქოლესტერინის ფონდის ფორმირების წყაროები და მისი ხარჯვის გზები მოცემულია ნახაზზე 22.1. სურ. 22.1. ორგანიზმში ქოლესტერინის წარმოქმნა და განაწილება. ადამიანის სხეულის ქოლესტერინი (საერთო რაოდენობა დაახლოებით 140 გ) პირობითად შეიძლება დაიყოს სამ აუზად: 30 გ), სწრაფად გაცვლისას, შედგება ნაწლავის კედლის ქოლესტერინის, სისხლის პლაზმის, ღვიძლის და სხვა პარენქიმული ორგანოების ქოლესტერინისგან, განახლება ხდება 30 დღეში (1 გ დღეში), 50 გ) სხვა ორგანოებისა და ქსოვილების ქოლესტერინის ნელა გაცვლით, 60 გ), ზურგის ტვინის და ტვინის ძალიან ნელა გაცვლილი ქოლესტერინი, შემაერთებელი ქსოვილი, განახლების სიჩქარე გამოითვლება წლების განმავლობაში. ქოლესტერინის სინთეზი ხდება უჯრედების ციტოზოლში. ეს არის ერთ – ერთი ყველაზე გრძელი მეტაბოლური გზა ადამიანის სხეულში. იგი მიმდინარეობს 3 სტადიაში: პირველი მთავრდება მევალონიუმის მჟავას წარმოქმნით, მეორე - სკალოლის წარმოქმნით (ხაზოვანი ნახშირწყალბადის სტრუქტურა, რომელიც შედგება 30 ნახშირბადის ატომისაგან). მესამე ეტაპის დროს, სკალალენი გარდაიქმნება ლანოსტერილის მოლეკულაში, შემდეგში არსებობს 20 თანმიმდევრული რეაქცია, რომლებიც ლანოსტერიოლს ქოლესტერინად გარდაქმნის. ზოგიერთ ქსოვილებში, ქოლესტერინის ჰიდროქსილიური ჯგუფი ასტერიზირებს ესთერების ფორმირებას. რეაქცია კატალიზირდება უჯრედშიდა ფერმენტის AHAT- ით (აცილქოა: ქოლესტერინის აცილტრანსფერაზა). ეთერიფიკაციის რეაქცია ასევე გვხვდება სისხლში HDL, სადაც მდებარეობს ფერმენტი LHAT (ლეციტინი: ქოლესტერინის აცილტრანსფერაზა). ქოლესტერინის ესტერია ის ფორმაა, რომლის საშუალებითაც იგი სისხლით ტრანსპორტირდება ან უჯრედებში იდება. სისხლში ქოლესტერინის დაახლოებით 75% ესთერების ფორმაა. ქოლესტერინის სინთეზი რეგულირდება პროცესის საკვანძო ფერმენტის აქტივობასა და რაოდენობაზე გავლენას ახდენს - 3-ჰიდროქსი-3-მეთილგლუტარილ-CoA რედუქტაზა (HMG-CoA რედუქტაზა). ეს მიიღწევა ორი გზით: 1. HMG-CoA რედუქტაზას ფოსფორილირება / დეფოსფორილიაცია. ინსულინი ასტიმულირებს HMG-CoA რედუქტაზას დეფოსფორილირებას, რითაც ის გადააქვთ აქტიურ მდგომარეობაში. ამიტომ, შთანთქმის პერიოდში, ქოლესტერინის სინთეზი იზრდება. ამ პერიოდის განმავლობაში, ასევე იზრდება საწყისი სუბსტრატის არსებობა სინთეზისთვის, აცეტილ- CoA– სთვის. გლუკაგონს აქვს საპირისპირო ეფექტი: A ცილის კინაზის საშუალებით იგი ასტიმულირებს HMG-CoA რედუქტაზას ფოსფორილირებას, გარდაქმნის მას არააქტიურ მდგომარეობაში. შედეგად, პოსტბსორბციის პერიოდში და მარხვის პერიოდში ქოლესტერინის სინთეზი შეფერხებულია. 2. HMG-CoA რედუქტაზას სინთეზის ინჰიბირება.ქოლესტერინი (მეტაბოლური ბილიკის საბოლოო პროდუქტი) ამცირებს HMG-CoA რედუქტაზის გენის ტრანსკრიპციის სიჩქარეს, რითაც შეაჩერებს საკუთარ სინთეზს, და ნაღვლის მჟავებიც იწვევს მსგავსი ეფექტს. სისხლის ქოლესტერინის ტრანსპორტირება ხორციელდება როგორც პრეპარატის ნაწილი. LPs უზრუნველყოფს ეგზოგენურ ქოლესტერინს ქსოვილებში, განსაზღვრავს მის ნაკადებს ორგანოებსა და ორგანიზმიდან ექსკრეციას. ეგზოგენური ქოლესტერინი ღვიძლს მიეწოდება, როგორც ნარჩენი ChM- ის ნაწილი. იქ სინთეზირებულ ენდოგენურ ქოლესტერინთან ერთად ქმნის საერთო ფონდს. ჰეპატოციტებში, TAGs და ქოლესტერინი შეფუთულია VLDL- ში და ამ ფორმით სისხლში სეკრეტირდება. სისხლში, VLDL LP- ლიპაზის მოქმედების ქვეშ, რომელიც TAG- ს ატარებს გლიცეროლსა და ცხიმოვან მჟავებს, პირველად გადააქვთ LSPP- ში, შემდეგ კი LDL- ში, რომელიც შეიცავს 55% ქოლესტერინს და მის ეთერებს. LDL არის ქოლესტერინის მთავარი სატრანსპორტო ფორმა, რომელშიც იგი მიეწოდება ქსოვილებს (ქოლესტერინის 70% და სისხლში მისი ესტერია LDL- ის ნაწილია). სისხლიდან LDL შედის ღვიძლში (75% -მდე) და სხვა ქსოვილებში, რომლებსაც აქვთ ზედაპირზე LDL რეცეპტორები. თუ უჯრედში შემავალი ქოლესტერინის რაოდენობა აღემატება მის საჭიროებას, თრგუნავს LDL რეცეპტორების სინთეზს, რაც ამცირებს სისხლიდან ქოლესტერინის გადინებას. უჯრედში თავისუფალი ქოლესტერინის კონცენტრაციის დაქვეითებით, პირიქით, გააქტიურებულია რეცეპტორების სინთეზი. ჰორმონები მონაწილეობენ LDL რეცეპტორების სინთეზის რეგულირებაში: ინსულინი, ტრიიოდთირონინი და სქესის ჰორმონები ზრდის რეცეპტორების წარმოქმნას, ხოლო გლუკოკორტიკოიდები მცირდება. ე.წ. "ქოლესტერინის დაბრუნების ტრანსპორტში", ე.ი. გზა, რომელიც უზრუნველყოფს ღვიძლის ქოლესტერინის დაბრუნებას, HDL მთავარ როლს ასრულებს. ისინი ღვიძლში სინთეზირდებიან გაუაზრებელი წინამორბედების სახით, რომლებიც პრაქტიკულად არ შეიცავს ქოლესტერინს და TAG- ს. სისხლში HDL წინამორბედები გაჯერებულია ქოლესტერინით, იღებენ მას სხვა LP– ებისა და უჯრედის მემბრანებისგან. ქოლესტერინის HDL– ში გადატანა მოიცავს მათ ზედაპირზე მდებარე LHAT ფერმენტს. ეს ფერმენტი ახდენს ცხიმოვანი მჟავების ნარჩენს ფოსფატიდილქოლინიდან (ლეციტინი) ქოლესტერინამდე. შედეგად, იქმნება ქოლესტერინის ეთერის ჰიდროფობიური მოლეკულა, რომელიც მოძრაობს HDL- ს შიგნით. ამრიგად, არ არის მთვრალი HDL, გამდიდრებული ქოლესტერინით, გადაიქცევა HDL 3-ში - მომწიფებული და უფრო დიდი ნაწილაკები. HDL 3 გაცვლით ქოლესტერინის ეთერებს TAG შეიცავს VLDL და STD სპეციფიკური ცილის მონაწილეობით, რომელიც გადადის ქოლესტერინის ესტრებს ლიპოპროტეინებს შორის. ამ შემთხვევაში, HDL 3 გადაიქცევა HDL2- ში, რომლის ზომა იზრდება TAG- ის დაგროვების გამო. VLDL და STDL LP- ლიპაზის გავლენის ქვეშ გარდაიქმნება LDL, რომლებიც ძირითადად ღვიძლში ქოლესტერინს მიაწვდიან. ქოლესტერინის მცირე ნაწილი მიეწოდება ღვიძლში HDL2 და HDL. ნაღვლის მჟავების სინთეზი. ღვიძლში, დღეში 5-700 მგ ნაღვლის მჟავების სინთეზი ხდება ქოლესტერინისგან. მათი ფორმირება მოიცავს ჰიდროქსილის ჯგუფების შეყვანის რეაქციებს ჰიდროქსილაზების მონაწილეობით და ქოლესტერინის გვერდითი ჯაჭვის ნაწილობრივი დაჟანგვის რეაქციაზე (ნახ. 22.2): სურ. 22.2. ნაღვლის მჟავების წარმოქმნის სქემა. პირველი სინთეზური რეაქცია - 7-ა-ჰიდროქსიქოლესტერინის წარმოქმნა - რეგულირებადია. ფერმენტის მოქმედება, რომელიც ახდენს ამ რეაქციის კატალიზაციას, ინჰიბირებულია გზის მჟავას საბოლოო პროდუქტით, ნაღვლის მჟავებით. რეგულირების კიდევ ერთი მექანიზმია ფერმენტის ფოსფორილირება / დეფოსფორილირება (7-ა-ჰიდროქსილაზას ფოსფორილირებული ფორმა აქტიურია). რეგულირება ასევე შესაძლებელია ფერმენტის ოდენობის შეცვლით: ქოლესტერინი იწვევს 7-ა-ჰიდროქსილაზა გენის ტრანსკრიპციას და ნაღვლის მჟავების რეპრესირებას. ფარისებრი ჯირკვლის ჰორმონები იწვევს 7-ა-ჰიდროქსილაზას სინთეზს და ესტროგენის რეპრესირებას. ესტროგენის ასეთი მოქმედება ნაღვლის მჟავების სინთეზზე ხსნის, თუ რატომ ხდება ნაღვლის ბუშტის დაავადება ქალებში 3-4 ჯერ უფრო ხშირად, ვიდრე მამაკაცებში. ქოლესტერინისგან წარმოქმნილ ქოლეს და ცენოდოქსიქოლულ მჟავებს უწოდებენ "პირველადი ნაღვლის მჟავას". ამ მჟავების უმეტესი ნაწილი გადის კონიუგაციას - გლიცინის ან ტაურინის მოლეკულების დამატება ნაღვლის მჟავის კარბოქსილის ჯგუფში. კონიუგაცია იწყება ნაღვლის მჟავების აქტიური ფორმის წარმოქმნით - CoA- ის წარმოებულები, შემდეგ თან ერთვის ტაურინი ან გლიცინი, რის შედეგადაც წარმოიქმნება კონიუგატების 4 ვარიანტი: ტაოროზული და ტაოროენოდოქსიქოლიური, გლიკოქოლური და გლიკოენოდოქსიქოლური მჟავები. ისინი მნიშვნელოვნად ძლიერი ემულგატორები არიან, ვიდრე თავდაპირველი ნაღვლის მჟავები. გლიცინთან ერთად კონიუგურები წარმოიქმნება 3 ჯერ უფრო მეტს, ვიდრე ტაურინთან, რადგან სხეულში ტაურინის რაოდენობა შეზღუდულია. ნაწლავში, მცირე რაოდენობით პირველადი ნაღვლის მჟავების კონიუგატები ბაქტერიული ფერმენტების მოქმედებით, გარდაიქმნება მეორად ნაღვლის მჟავებად. დეოქსიქოლური მჟავა, რომელიც წარმოქმნილი ქოქოლიდან და ლითოქოლურიდან, ჩამოყალიბებულია დეოქსიქოლურიდან, ნაკლებად ხსნადი და უფრო ნელა შეიწოვება ნაწლავებში. ნაღვლის მჟავების 95%, რომლებიც ნაწლავში შედიან, ღვიძლში ღვიძლში უბრუნდება პორტალური ვენის საშუალებით, შემდეგ კვლავ გამოიყოფა ნაღვლის ღრუში და მეორდება ცხიმების ემულსიფიკაციაში. ნაღვლის მჟავების ამ გზას ეწოდება ენტეროჰეპტიკური მიმოქცევა. განავლით, მეორადი ნაღვლის მჟავები ძირითადად ამოღებულია. ნაღვლის ბუშტის დაავადება (ქოლელთიოზი) არის პათოლოგიური პროცესი, რომლის დროსაც ქვები ვითარდება ნაღვლის ბუშტში, რომლის საფუძველია ქოლესტერინი. ქოლესტერინის ნაღველში მოთავსებას თან უნდა ახლდეს ნაღვლის მჟავების და ფოსფოლიპიდების პროპორციული განთავისუფლება, რომლებიც ინარჩუნებენ ჰიდროფობიური ქოლესტერინის მოლეკულებს მიკელარულ მდგომარეობაში. ნაღვლის მჟავების და ქოლესტერინის თანაფარდობის ცვლილების მიზეზები არის: ქოლესტერინით მდიდარი საკვები, მაღალკალორიული კვება, ნაღვლის ბუშტში ნაღვლის სტაგნაცია, ენტეროჰეპტიკური მიმოქცევის დაქვეითება, ნაღვლის მჟავების სინთეზის დაქვეითება, ნაღვლის ბუშტის ინფექცია. ქოლელითაზიის მქონე პაციენტთა უმრავლესობაში იზრდება ქოლესტერინის სინთეზი და ნელდება მისგან ნაღვლის მჟავების სინთეზი, რაც იწვევს დისპორციულობას ქოლესტერინის და ნაღვლის მჟავების სეკრეციის დროს. შედეგად, ქოლესტერინი იწყებს ნალექს ნაღვლის ბუშტში, ქმნის ბლანტი ნალექს, რომელიც თანდათან ძლიერდება. ზოგჯერ გაჟღენთილია ბილირუბინი, ცილები და კალციუმის მარილები. ქვები შეიძლება შედგებოდეს მხოლოდ ქოლესტერინისგან (ქოლესტერინის ქვები) ან ქოლესტერინის, ბილირუბინის, ცილებისა და კალციუმის ნაზავი. ქოლესტერინის ქვები, როგორც წესი, თეთრია, ხოლო შერეული ქვები ყავისფერია სხვადასხვა ფერებში. ქვის წარმოქმნის საწყის ეტაპზე, ფენოქსიქსიქოლური მჟავა შეიძლება გამოყენებულ იქნას, როგორც წამალი. ნაღვლის ბუშტში ყოფნის შემდეგ იგი თანდათანობით დაითხოვს ქოლესტერინის ქვებს, მაგრამ ეს ნელი პროცესია, რომელიც გრძელდება რამდენიმე თვეს. ათეროსკლეროზი არის პათოლოგია, რომელიც ხასიათდება სისხლძარღვთა კედლის შიდა ზედაპირზე ათეროგენული დაფების გამოჩენით. ამგვარი პათოლოგიის განვითარების ერთ-ერთი მთავარი მიზეზი არის საკვებიდან ქოლესტერინის მიღებას შორის ბალანსის დარღვევა, მისი სინთეზი და ორგანიზმიდან გამოდევნა. ათეროსკლეროზის მქონე პაციენტებში აღინიშნება LDL და VLDL კონცენტრაცია. არსებობს HDL კონცენტრაციასთან და ათეროსკლეროზის განვითარების ალბათობას შორის საპირისპირო ურთიერთობა. ეს შეესაბამება LDL- ს ფუნქციონირების კონცეფციას, როგორც ქსოვილებში ქოლესტერინის მატარებლები, ხოლო ქსოვილებისგან HDL. ათეროსკლეროზის განვითარების ძირითადი მეტაბოლური "წინაპირობა" არის ჰიპერქოლესტერინემია. (მაღალი ქოლესტერინი სისხლში). ვითარდება ჰიპერქოლესტერინემია: 1. ქოლესტერინის, ნახშირწყლების და ცხიმების ჭარბი მიღების გამო, 2. გენეტიკური მიდრეკილება, რომელიც შედგება მემკვიდრეობითი დეფექტების შემადგენლობაში LDL ან apoB-100 რეცეპტორების სტრუქტურაში, აგრეთვე ა apoB-100 სეკრეციის გაზრდის დროს (ოჯახური კომბინირებული ჰიპერლიპიდემიის შემთხვევაში, რომელშიც აღინიშნება სისხლის კონცენტრაცია და ქოლესტერინი და TAG). ათეროსკლეროზის განვითარების მექანიზმებში მნიშვნელოვან როლს ასრულებს პრეპარატის მოდიფიცირება. LDL- ში ლიპიდების და ცილების ნორმალურ სტრუქტურაში ცვლილებები მათ უცხო ორგანიზმს უქმნის და, შესაბამისად, უფრო ხელმისაწვდომი გახდება ფაგოციტების მიერ. წამლის მოდიფიცირება შეიძლება მოხდეს რამდენიმე მექანიზმით: 1. ცილების გლიკოზილაცია, რომელიც ხდება სისხლში გლუკოზის კონცენტრაციის გაზრდის დროს, 2. პეროქსიდის მოდიფიკაცია, რამაც გამოიწვია ლიპოპროტეინებში ლიპიდების ცვლილებები და apoB-100- ის სტრუქტურა, 3. LP- ანტისხეულების აუტოიმუნური კომპლექსების წარმოქმნა (შეცვლილმამა წამლებმა შეიძლება გამოიწვიოს აუტოანტისხეულების წარმოქმნა). მოდიფიცირებული LDL შეიწოვება მაკროფაგებით. ეს პროცესი არ რეგულირდება აბსორბირებული ქოლესტერინის ოდენობით, ისევე როგორც მისი სპეციფიკური რეცეპტორების საშუალებით უჯრედებში მისი შეყვანის შემთხვევაში, ამიტომ მაკროფაგები გადატვირთულია ქოლესტერინით და გადაიქცევა „ქაფებად უჯრედებად“, რომლებიც შედიან სუბენდოთელურ სივრცეში. ეს იწვევს სისხლძარღვების კედელში ლიპიდური ლაქების ან ზოლების წარმოქმნას. ამ ეტაპზე სისხლძარღვთა ენდოთელიუმს შეუძლია შეინარჩუნოს მისი სტრუქტურა. ქაფის უჯრედების რაოდენობის მატებასთან ერთად ენდოთელური დაზიანება ხდება. დაზიანება ხელს უწყობს თრომბოციტების გააქტიურებას. შედეგად, ისინი ასუფთავებენ თრომბოქსანს, რომელიც ასტიმულირებს თრომბოციტების აგრეგაციას და ასევე იწყებენ თრომბოციტების ზრდის ფაქტორს, რაც ასტიმულირებს გლუვი კუნთების უჯრედების გამრავლებას. ეს უკანასკნელი მედიკაციიდან მიემართება არტერიული კედლის შიდა შრეზე, რითაც ხელს უწყობს დაფის ზრდას. გარდა ამისა, ბოჭკოვანი ქსოვილისგან დაფა დაფარულია, ბოჭკოვანი გარსის ქვეშ მყოფი უჯრედები ნეკროზულია, ქოლესტერინი კი დეპონირდება უჯრედშორის სივრცეში. განვითარების ბოლო ეტაპზე, დაფა გაჟღენთილია კალციუმის მარილებით და ხდება ძალიან მკვრივი. დაფის მიდამოში ხშირად სისხლის კოლტები წარმოიქმნება, გემის სანათურის ბლოკირება, რაც იწვევს ქსოვილების მწვავე უკმარისობას შესაბამის ქსოვილების ადგილზე და გულის შეტევის განვითარებას. ქოლესტერინის სინთეზის რეგულირება - მისი საკვანძო ფერმენტი (HMG-CoA რედუქტაზა) ხორციელდება სხვადასხვა გზით. HMG რედუქტაზას ფოსფორილირება / დეფოსფორილირება. ინსულინ / გლუკაგონის თანაფარდობის მატებით, ეს ფერმენტი ფოსფორილდება და გადადის აქტიურ მდგომარეობაში. ინსულინის მოქმედება ხორციელდება 2 ფერმენტის მეშვეობით. HMG-CoA ამცირებს კინაზას ფოსფატაზას, რომელიც კინაზას გადააქცევს არააქტიურ დეფოსფორილირებულ მდგომარეობაში: ფოსფოტაზა HMG-CoA რედუქტაზა მისი დეფოსფორილირებული აქტიურ მდგომარეობაში გადაკეთებით. ამ რეაქციების შედეგია HMG-CoA რედუქტაზის დეფოსფორილირებული აქტიური ფორმის წარმოქმნა. შესაბამისად, შთანთქმის პერიოდში, ქოლესტერინი იზრდება. ამ პერიოდის განმავლობაში, საწყისი სუბსტრატის არსებობა ხდება ქოლესტერინის - აცეტილ - CoA სინთეზისთვის (ნახშირწყლებისა და ცხიმების შემცველი საკვების ჭამის შედეგად, რადგან გლუკოზისა და ცხიმოვანი მჟავების დაშლის დროს წარმოიქმნება აცეტილ CoA). პოსტაბსორბენტულ მდგომარეობაში გლუკაგონი A პროტეგენგენაზას საშუალებით ასტიმულირებს HMG - CoA - ფოსფორილირებას - რედუქტაზას, გადააქცევს მას არააქტიურ მდგომარეობაში. ეს მოქმედება გაძლიერებულია იმით, რომ ამავე დროს გლუკაგონი ასტიმულირებს HMG-CoA რედუქტაზას ფოსფოტაზას ფოსფორილირებას და ინაქტივირებას, რითაც HMG-CoA რედუქტაზას ინარჩუნებს ფოსფორილირებულ არააქტიურ მდგომარეობაში. შედეგად, პოსტბსორბციის პერიოდში და მარხვის პერიოდში ქოლესტერინის სინთეზი ინჰიბირდება ენდოგენური სინთეზით. თუ საკვებში ქოლესტერინის შემცველობა 2% -მდე იქნა მიღებული, მაშინ ენდოგენური ქოლესტერინის სინთეზი მკვეთრად შემცირდა. მაგრამ ქოლესტერინის სინთეზის სრული შეწყვეტა არ ხდება. საკვებიდან მომავალი ქოლესტერინის გავლენის ქვეშ ქოლესტერინის ბიოსინთეზის ინჰიბირების ხარისხი განსხვავდება პიროვნებიდან. ეს მიუთითებს ქოლესტერინის წარმოქმნის პროცესების ინდივიდუალურობაზე. ქოლესტერინის სინთეზის ინტენსივობის შემცირებით შესაძლებელია სისხლში მისი კონცენტრაციის შემცირება. თუ ერთის მხრივ ორგანიზმში ქოლესტერინის საკვების მიღებასა და მასში სინთეზს შორის წონასწორობა და მეორეს მხრივ, ნაღვლის მჟავების და ქოლესტერინის ექსკრეცია დარღვეულია, ქსოვილებში ქოლესტერინის კონცენტრაცია და სისხლი იცვლება. ყველაზე სერიოზული შედეგები უკავშირდება სისხლში ქოლესტერინის კონცენტრაციის ზრდას (ჰიპერქოლესტერინემია), ხოლო ათეროსკლეროზისა და ქოლელითაზიის განვითარების ალბათობა იზრდება. ოჯახური ჰიპერქოლესტერინემია (HCS) - ეს ფორმა გაცილებით გავრცელებულია - 200 ადამიანზე დაახლოებით 1 პაციენტი. HCS- ში მემკვიდრეობითი დეფექტი არის უჯრედების მიერ LDL შეწოვის დარღვევა და, შესაბამისად, LDL კატაბოლიზმის სიჩქარის დაქვეითება. შედეგად, სისხლში LDL- ის კონცენტრაცია იზრდება, ისევე როგორც ქოლესტერინი, რადგან მასში ბევრია LDL- ში. ამიტომ, HCS– ით, ქოლესტერინის დეპონირება ხდება ქსოვილებში, კერძოდ კანში (ქსანთომა), არტერიების კედლებში. HMG-CoA რედუქტაზას სინთეზის ინჰიბირება ქოლესტერინის მეტაბოლური გზის საბოლოო პროდუქტი. ეს ამცირებს HMG-CoA რედუქტაზას გენის ტრანსკრიპციის სიჩქარეს, რითაც ახდენს საკუთარი სინთეზის შეფერხებას. ღვიძლი აქტიურად ახდენს ნაღვლის მჟავების სინთეზს ქოლესტერინისგან, და ამიტომ ნაღვლის მჟავები აფერხებენ HMG-CoA რედუქტაზის გენის მოქმედებას. ვინაიდან HMG-CoA რედუქტაზა არსებობს დაახლოებით 3-ის სინთეზის შემდეგ, ამ ქოლესტერინის ფერმენტის სინთეზის დათრგუნვა ეფექტური რეგულირებაა.
|